Adoptive T-cell therapies rely on generating metabolically fit, functional cells during ex vivo expansion—but this process often pushes T cells toward highly glycolytic, terminally differentiated states that limit their persistence and therapeutic potential. These metabolic programs begin shifting within hours of activation, therefore understanding early metabolic remodeling is essential for designing culture conditions that support durable, cytotoxic, and memory-enriched T-cell populations.
Researchers at Promega set out to address this challenge by systematically mapping how media composition and activation strength shape T-cell metabolism during the first 72 hours after stimulation. Using a suite of bioluminescent assays, they profiled intracellular energy cofactors, redox balance, and extracellular metabolites across several conditions. This approach revealed distinct, media-driven metabolic states that not only emerged early but also predicted downstream expansion, proliferation, and cytotoxic function.
Their work demonstrates how integrating metabolic profiling into in vitro expansion workflows can provide a more informed framework for optimizing T-cell manufacturing strategies.
Attention-Deficit/Hyperactivity Disorder (ADHD) is a complex neurodevelopmental disorder that affects millions worldwide. Current therapeutic treatment relies on pharmaceutical approaches, but emerging research suggests that dietary supplements, such as omega-3 fatty acids, may offer complementary therapeutic options. A recent study published in the Journal of Psychiatric Research explores the relationship between inflammation and dietary supplements to determine how they might influence ADHD pathology. This work was conducted in Dr. Edna Grünblatt’s lab at the University of Zurich and was supported through Promega’s Academic Access Program. I had the chance to interview Dr. Natalie Walter, the lead author, to learn more about how her work offers potential opportunities for non-pharmacological interventions.
It’s been just over 10 years since the world lost a pioneering immunologist and biochemist, Dr. Jürg Tschopp. He died tragically during a hiking trip in the Swiss Alps on March 22, 2011. A host of academic journals, including Science, Nature and Cell, paid tribute to Dr. Tschopp with obituaries that highlighted his many accomplishments in the fields of apoptosis and immunology.
In 2002, a team led by Dr. Tschopp at the University of Lausanne, Switzerland, was studying the role of the proinflammatory cytokine interleukin 1 beta (IL-1β). This cytokine is produced in the cytoplasm as an inactive precursor (pro-IL-1β). It is cleaved by caspase-1 to the active form, but the exact process by which caspase-1 itself is activated was unknown at the time. Several members of the caspase family contain a conserved region known as the caspase recruitment domain or CARD, and it was proposed that this domain was essential to caspase activation.
Based on similarity to another protein containing an N-terminal CARD motif (Apaf-1) that is involved in activation of caspase-9, the researchers examined the roles of a family of proteins known as NALP1, NALP2 and NALP3 (1). In particular, they were interested in NALP1, which is involved in the immune response. Unlike Apaf-1, NALP1 contains a CARD motif at the C terminus, while the N terminus contains a related motif known as a pyrin-like domain (PYD). The research team had previously showed that the PYD region of NALP1 interacted with an adapter protein known as PYCARD or ASC, which also contains an N-terminal PYD and C-terminal CARD.
The results of the team’s in vitro binding, activation and immunodetection studies showed that a multi-unit protein complex is responsible for caspase activation, and they called this complex the “inflammasome” (1). It is composed of caspase-1, caspase-5, PYCARD/ASC and NALP1.
Today’s blog is written by the University of Copenhagen iGEM Team.
The International Genetically Engineered Machine (iGEM) competition has 257 teams of students competing this year. Despite all of the unique difficulties we’re all facing in 2020, the University of Copenhagen is competing once again. This year’s project involves a unique approach to Chronic Inflammatory Diseases (CIDs).
Our innate immune system was meant to do good. Up until a
century ago, most humans died from infectious diseases like diarrhea,
tuberculosis and meningitis. Over millions of years, our immune system has
evolved to fight these life-threatening infections from pathogens. As a result,
we have developed a highly efficient response to these tiny invaders. But it
seems that our immune system may be turning against us.
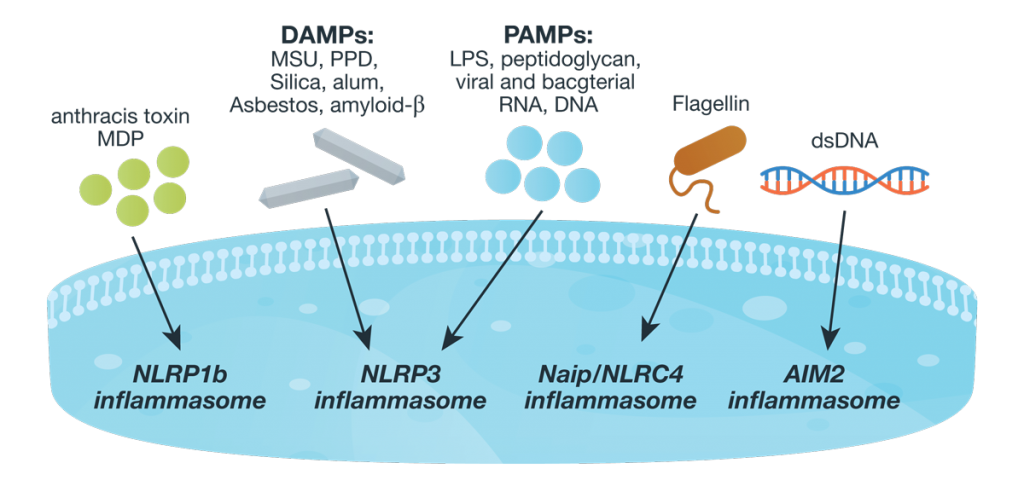
Innate immunity, the first line of immune defense, uses a system of host pattern recognition receptors (PRRs) to recognize signals of “danger” including invariant pathogen-associated molecular patterns (PAMPs) and damage-associated molecular patterns (DAMPs). These signals in turn recruit and assemble protein complexes called inflammasomes, resulting in the activation of caspase-1, the processing and release of the pro-inflammatory cytokines IL-1ß and IL-18, and the induction of programmed, lytic cell death known as pyroptosis.
Innate immunity and the activity of the inflammasome are critical for successful immunity against a myriad of environmental pathogens. However dysregulation of inflammasome activity is associated with many inflammatory diseases including type 2 diabetes, obesity-induced asthma, and insulin resistance. Recently, aberrant NLRP3 inflammasome activity also has been associated with age-related macular degeneration and Alzheimer disease. Understanding the players and regulators involved in inflammasome activity and regulation may provide additional therapeutic targets for these diseases.
Currently inflammasome activation is monitored using antibody-based techniques such as Western blotting or ELISA’s to detect processed caspase-1 or processed IL-1ß. These techniques are tedious and are only indirect measures of caspase activity. Further, gaining information about kinetics—relating inflammasome assembly, caspase-1 activation and pyroptosis in time—is very difficult using these methods. O’Brien et al. describe a one-step, high-throughput method that enables the direct measurement of caspase-1 activity. The assay can be multiplexed with a fluorescent viability assay, providing information about the timing of cell death and caspase-1 activity from the same sample. Continue reading “Activating the Inflammasome: A New Tool Brings New Understanding”
In today’s post, guest blogger, Martha O’Brien, PhD, provides a preview of her upcoming AAI poster and block symposium talk on the inflammasome, caspase-1 activity and pyroptosis.
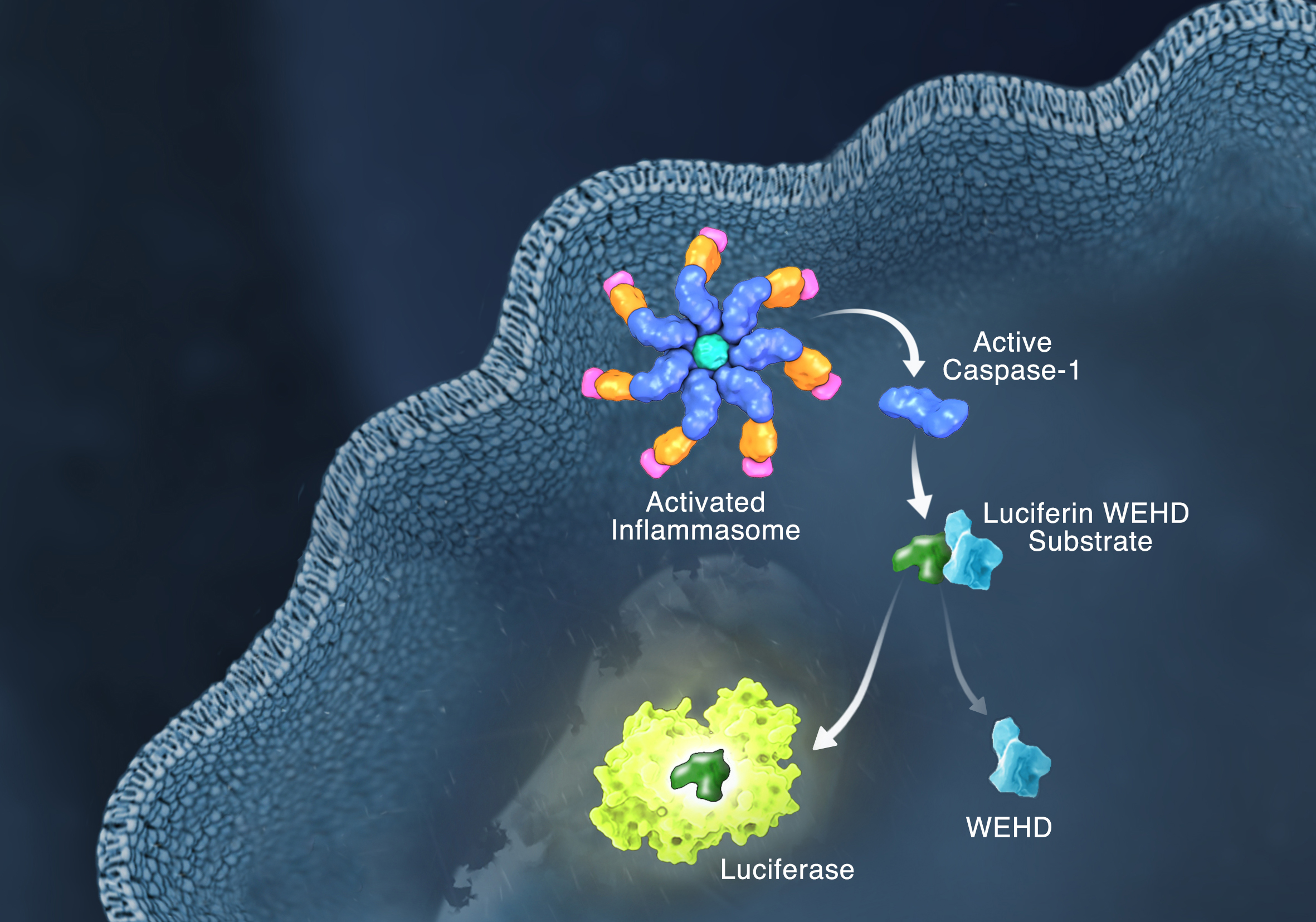
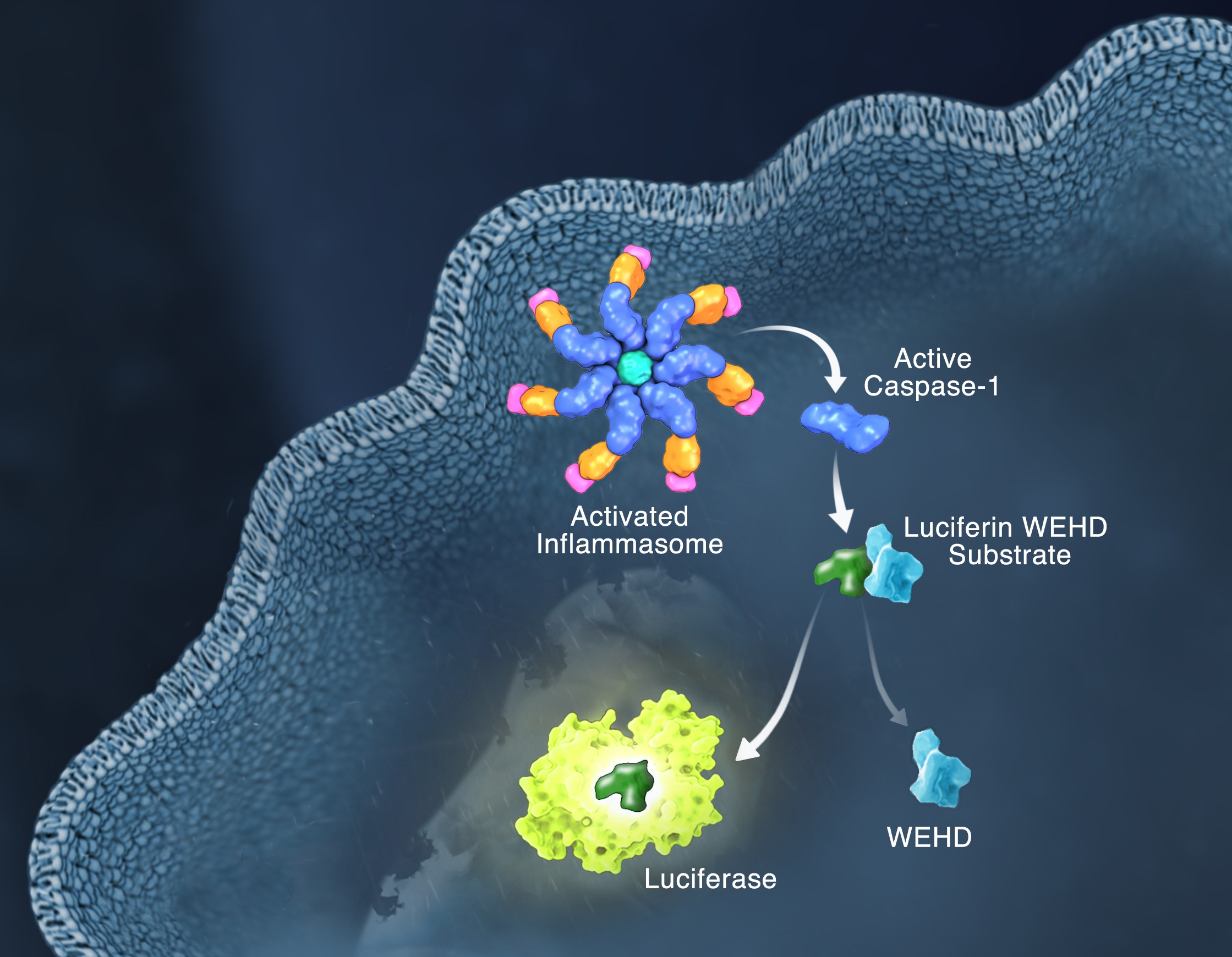
Schematic of the Caspase-Glo 1 Inflammasome Assay.
Responding rapidly to microbial pathogens and damage-associated molecular markers is critical to our innate immune system. Caspase-1 is pivotal in this process leading to processing and release of essential cytokines and an immunogenic form of cell death, termed pyroptosis. Upon sensing pathogen-associated and damage-associated molecular patterns (PAMPs and DAMPs), innate immune cells form inflammasome protein complexes that recruit and activate caspase-1 (canonical inflammasomes). In addition, other inflammatory caspases, 4 and 5 in humans and 11 in mice, directly bind bacterial lipopolysaccharides (LPS), triggering pyroptosis (non-canonical inflammasome). LPS-triggered non-canonical inflammasomes in mice and humans ultimately lead to canonical inflammasome engagement and caspase-1 activation (1–3). Caspase-1 was originally termed interleukin converting enzyme (ICE) for its well-established role in processing IL-1ß and IL-18, two important inflammation cytokines. How caspase-1 mediates pyroptosis is less well understood, but is beginning to be delineated. Recently, a substrate of the inflammatory caspases, gasdermin D, was identified and its processed fragment, gasdermin-N domain, was shown to be required for pyroptosis in non-canonical inflammasome circumstances (4, 5). The precise role of gasdermin D in canonical inflammasome-triggered pyroptosis is still under investigation. Linking inflammatory caspases directly to pyroptosis is a notable step in understanding the mechanism of this important form of cell death.
Pyroptosis is clearly one means of releasing processed IL-1ß and IL-18 from the cell. However depending on the cell type and stimulus, there is evidence for inflammasome engagement, caspase-1 activation, and release of IL-1ß in the absence of cell death (6, 7). On the flip-side there is also evidence for caspase-1 mediated pyroptosis that helps clear bacteria, independent of IL-1ß and IL-18 involvement (8). To enable further studies on the inflammasome and in particular, assessing the connections between caspase-1 activation, pyroptosis, and cytokine release, Promega developed a new tool to conveniently monitor caspase-1 activation, the Caspase-Glo® 1 Inflammasome Assay. This bioluminescent, plate-based assay is used to measure caspase-1 activity directly in cell cultures or to monitor released caspase-1 activity in culture medium from treated cells. This flexibility allows easy multiplexing to monitor all three outcomes of inflammasome stimulation; caspase-1 activity, pyroptosis, and release of IL-1ß and IL-18. Caspase-1 activation typically is monitored indirectly with western blots of processed caspase-1. Now the activity of the enzyme can be monitored directly, providing accurate information on temporal aspects of the inflammasome. The assay can be readily combined with real-time measures of cell death (e.g., CellTox™ Green Cytotoxicity Assay) and some of the culture medium can be removed for IL-1ß/IL-18 assessment, leaving the cells and remaining culture medium for caspase-1 activity measurements.
Most of us have experienced an inflammatory response at some point in our lives. Fever, achy joints, swelling around a scrape or cut, all of these are forms of inflammatory response. Inflammation is the body’s response to infection or tissue damage and acts to limit harm to the rest of the body. A key player in the inflammation process is a group of protein complexes call inflammasomes. The term “inflammasome” was first used in 2002 by researchers in Switzerland (1) to refer to a caspase-activating protein complex. We now know that inflammasomes are cytosolic multiprotein platforms that assemble in response to pathogens and other signals. Inflammasome assembly results in the processing of the inactive procaspase-1 into the active cysteine-protease enzyme, caspase-1. Caspase-1, in turn, activates the proinflammatory cytokines Interleukins IL-1β and IL-18. In addition, caspase-1 is also required for pyroptosis, which is an inflammatory form of cell death that combines the characteristics of apoptosis (fragmented DNA) and necrosis (inflammation and cytokine release) and is frequently associated with microbial infections.
Inflammasome complexes are made up of scaffolding sensor proteins (NLR, AIM2, ALR), and an adaptor protein containing a caspase activation and retention domain (CARD) and inactive procaspase-1. Most inflammasomes are formed with one or two NLRs (NOD-like receptors). However, non-NLR proteins such as AIM2 (absent in melanoma 2) and pyrin can also form inflammasomes. The different sensor proteins are activated by different types of outside stimuli, and inflammasomes are loosely sorted into families based on the protein forming these sensors.Continue reading “Inflammasomes: Peeking Inside the Inflammatory Process”
Anyone who has travelled across time zones knows how unpleasant it is when the regular rhythm of your biological clock is disrupted. Jetlag results when the body’s internal clock, or circadian rhythm is out of sync with external cues for “day and “night”, resulting in insomnia, extreme tiredness, difficulty concentrating and various other unpleasant symptoms.
On the bright side, jetlag is at least a temporary misery that is usually over after a few days of acclimation to the new time zone. Long-term disruption of the natural sleep/wake cycle, such as encountered by frequent long-distance travellers, shift workers, or people with physiological conditions that affect circadian rhythms, can be much more debilitating. Longer term health effects that have been associated with constant disruption of circadian rhythms include, insomnia, concentration problems, and increased susceptibility to diseases associated with chronic inflammation such as cancer, diabetes and cardiovascular disease.
Despite the fact that many of the genes and proteins involved in central control of circadian rhythms are known, the reason for the implied association between circadian clock components and immune function is not understood. Recently, a paper was published in the July issue of PNAS that identified a potential link between a circadian clock component and chronic inflammation. Continue reading “Molecular Connections Between Sleep Deprivation and Inflammation”
The majority of the ground transporation in the United States runs on diesel fuel. It powers most of the fleet of large vehicles that travel around Wisconsin and across country moving products and people. My family uses diesel fuel in all the large tractors that plant and harvest the 1,200+ acres of crops on the farm as well in the semitrailer that hauls the final product to market. Diesel also fuels a small portion of the passenger vehicles that people drive every day in the United States, including the car that is my main mode of transport. However, exposure to diesel exhaust can have negative effects on human health, and recent reports have found an increased risk of lung cancer. While these studies documented heavy exposure levels (miners underground with diesel generators), people still wonder about the effect of exposure when behind a truck spewing dark exhaust. A recent PLoS ONE article examined the effect of diesel exhaust particles on mouse cardiovascular systems and how treating with curcumin attenuated the negative effects. Continue reading “Curcumin Moderates the Health Effects of Diesel Exhaust”
XWe use cookies and similar technologies to make our website work, run analytics, improve our website, and show you personalized content and advertising. Some of these cookies are essential for our website to work. For others, we won’t set them unless you accept them. To learn more about our approach to Privacy we invite you to Read More
By clicking “Accept All”, you consent to the use of ALL the cookies. However you may visit Cookie Settings to provide a controlled consent.
We use cookies and similar technologies to make our website work, run analytics, improve our website, and show you personalized content and advertising. Some of these cookies are essential for our website to work. For others, we won’t set them unless you accept them. To find out more about cookies and how to manage cookies, read our Cookie Policy.
If you are located in the EEA, the United Kingdom, or Switzerland, you can change your settings at any time by clicking Manage Cookie Consent in the footer of our website.
Necessary cookies are absolutely essential for the website to function properly. These cookies ensure basic functionalities and security features of the website, anonymously.
Cookie
Duration
Description
cookielawinfo-checbox-analytics
11 months
This cookie is set by GDPR Cookie Consent plugin. The cookie is used to store the user consent for the cookies in the category "Analytics".
cookielawinfo-checbox-functional
11 months
The cookie is set by GDPR cookie consent to record the user consent for the cookies in the category "Functional".
cookielawinfo-checbox-others
11 months
This cookie is set by GDPR Cookie Consent plugin. The cookie is used to store the user consent for the cookies in the category "Other.
cookielawinfo-checkbox-advertisement
1 year
The cookie is set by GDPR cookie consent to record the user consent for the cookies in the category "Advertisement".
cookielawinfo-checkbox-necessary
11 months
This cookie is set by GDPR Cookie Consent plugin. The cookies is used to store the user consent for the cookies in the category "Necessary".
cookielawinfo-checkbox-performance
11 months
This cookie is set by GDPR Cookie Consent plugin. The cookie is used to store the user consent for the cookies in the category "Performance".
gdpr_status
6 months 2 days
This cookie is set by the provider Media.net. This cookie is used to check the status whether the user has accepted the cookie consent box. It also helps in not showing the cookie consent box upon re-entry to the website.
lang
This cookie is used to store the language preferences of a user to serve up content in that stored language the next time user visit the website.
viewed_cookie_policy
11 months
The cookie is set by the GDPR Cookie Consent plugin and is used to store whether or not user has consented to the use of cookies. It does not store any personal data.
Analytical cookies are used to understand how visitors interact with the website. These cookies help provide information on metrics the number of visitors, bounce rate, traffic source, etc.
Cookie
Duration
Description
SC_ANALYTICS_GLOBAL_COOKIE
10 years
This cookie is associated with Sitecore content and personalization. This cookie is used to identify the repeat visit from a single user. Sitecore will send a persistent session cookie to the web client.
vuid
2 years
This domain of this cookie is owned by Vimeo. This cookie is used by vimeo to collect tracking information. It sets a unique ID to embed videos to the website.
WMF-Last-Access
1 month 18 hours 24 minutes
This cookie is used to calculate unique devices accessing the website.
_ga
2 years
This cookie is installed by Google Analytics. The cookie is used to calculate visitor, session, campaign data and keep track of site usage for the site's analytics report. The cookies store information anonymously and assign a randomly generated number to identify unique visitors.
_gid
1 day
This cookie is installed by Google Analytics. The cookie is used to store information of how visitors use a website and helps in creating an analytics report of how the website is doing. The data collected including the number visitors, the source where they have come from, and the pages visted in an anonymous form.
Advertisement cookies are used to provide visitors with relevant ads and marketing campaigns. These cookies track visitors across websites and collect information to provide customized ads.
Cookie
Duration
Description
IDE
1 year 24 days
Used by Google DoubleClick and stores information about how the user uses the website and any other advertisement before visiting the website. This is used to present users with ads that are relevant to them according to the user profile.
test_cookie
15 minutes
This cookie is set by doubleclick.net. The purpose of the cookie is to determine if the user's browser supports cookies.
VISITOR_INFO1_LIVE
5 months 27 days
This cookie is set by Youtube. Used to track the information of the embedded YouTube videos on a website.
Performance cookies are used to understand and analyze the key performance indexes of the website which helps in delivering a better user experience for the visitors.
Cookie
Duration
Description
YSC
session
This cookies is set by Youtube and is used to track the views of embedded videos.
_gat_UA-62336821-1
1 minute
This is a pattern type cookie set by Google Analytics, where the pattern element on the name contains the unique identity number of the account or website it relates to. It appears to be a variation of the _gat cookie which is used to limit the amount of data recorded by Google on high traffic volume websites.

 Anyone who has travelled across time zones knows how unpleasant it is when the regular rhythm of your biological clock is disrupted. Jetlag results when the body’s internal clock, or circadian rhythm is out of sync with external cues for “day and “night”, resulting in insomnia, extreme tiredness, difficulty concentrating and various other unpleasant symptoms.
Anyone who has travelled across time zones knows how unpleasant it is when the regular rhythm of your biological clock is disrupted. Jetlag results when the body’s internal clock, or circadian rhythm is out of sync with external cues for “day and “night”, resulting in insomnia, extreme tiredness, difficulty concentrating and various other unpleasant symptoms.