We made the cover! Of Cell Chemical Biology, that is.
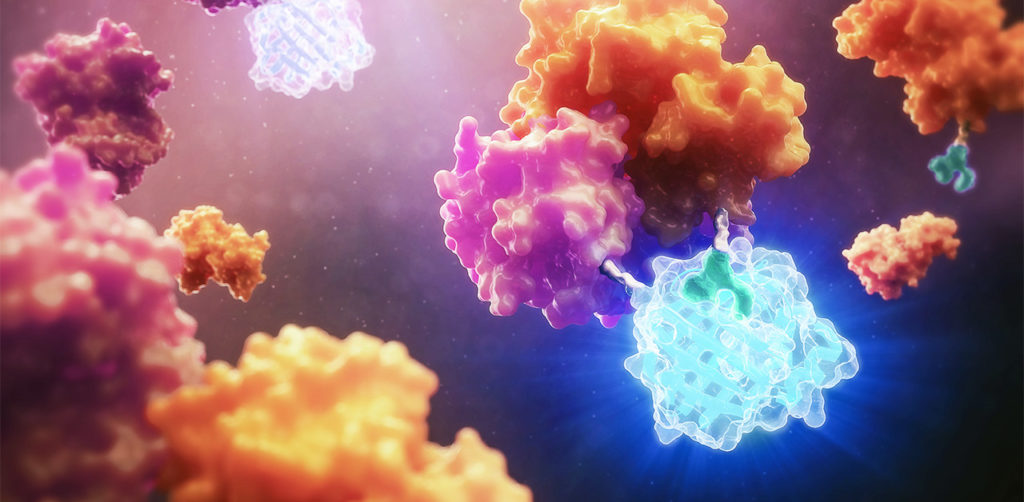
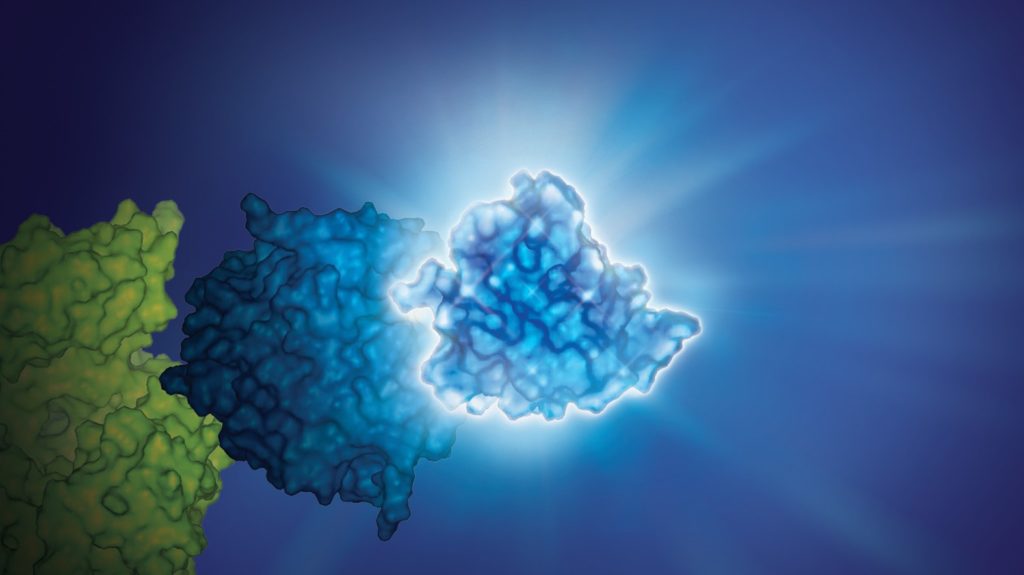
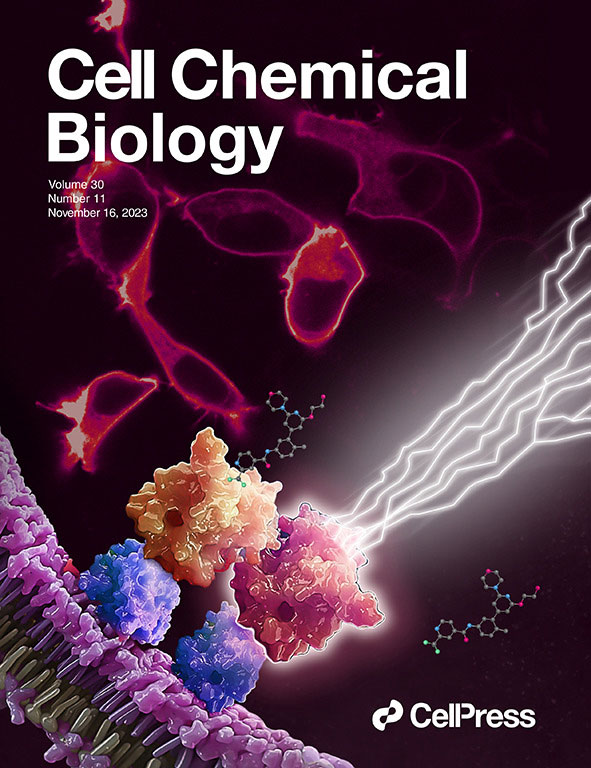
This July, Cell Chemical Biology editors accepted a study from Promega scientists and invited the research team to submit cover art for the issue. The study in question details a BRET-based method to quantify drug-target occupancy within RAF-KRAS complexes in live cells. Promega scientists Matt Robers and Jim Vasta collaborated with one of our talented designers, Michael Stormberg, to craft an image that accurately represents the science in a dynamic and engaging way.
You can check out the paper and cover art in the November 16 issue of Cell Chemical Biology.
I spoke with Michael Stormberg to learn more about the creative process that went into creating this cover art and how he worked with the research team and other collaborators.
Continue reading “Designing Science: A Behind-the-Scenes Look at Our Recent Journal Cover Art”