Understanding the expression, function and dynamics of proteins in their native environment is a fundamental goal that’s common to diverse aspects of molecular and cell biology. To study a protein, it must first be labeled—either directly or indirectly—with a “tag” that allows specific and sensitive detection.
Using a labeled antibody to the protein of interest is a common method to study native proteins. However, antibody-based assays, such as ELISAs and Western blots, are not suitable for use in live cells. These techniques are also limited by throughput and sensitivity. Further, suitable antibodies may not be available for the target protein of interest.
Continue reading “CRISPR/Cas9 Knock-In Tagging: Simplifying the Study of Endogenous Biology”



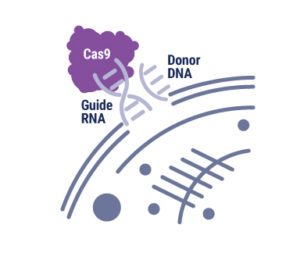
 GPCRs
GPCRs