At the time of writing this post, no scientist had yet discovered the secret to immortality. In our world, we’ve come to accept that living things are born, grow old and die—the circle of life.
And yet, for many years, life scientists believed that the circle of life did not apply to our constituent cells when cultured in a laboratory. That is, cultured normal human cells were immortal, and they would continue to grow and proliferate forever, as long as they were provided with the necessary nutrients.
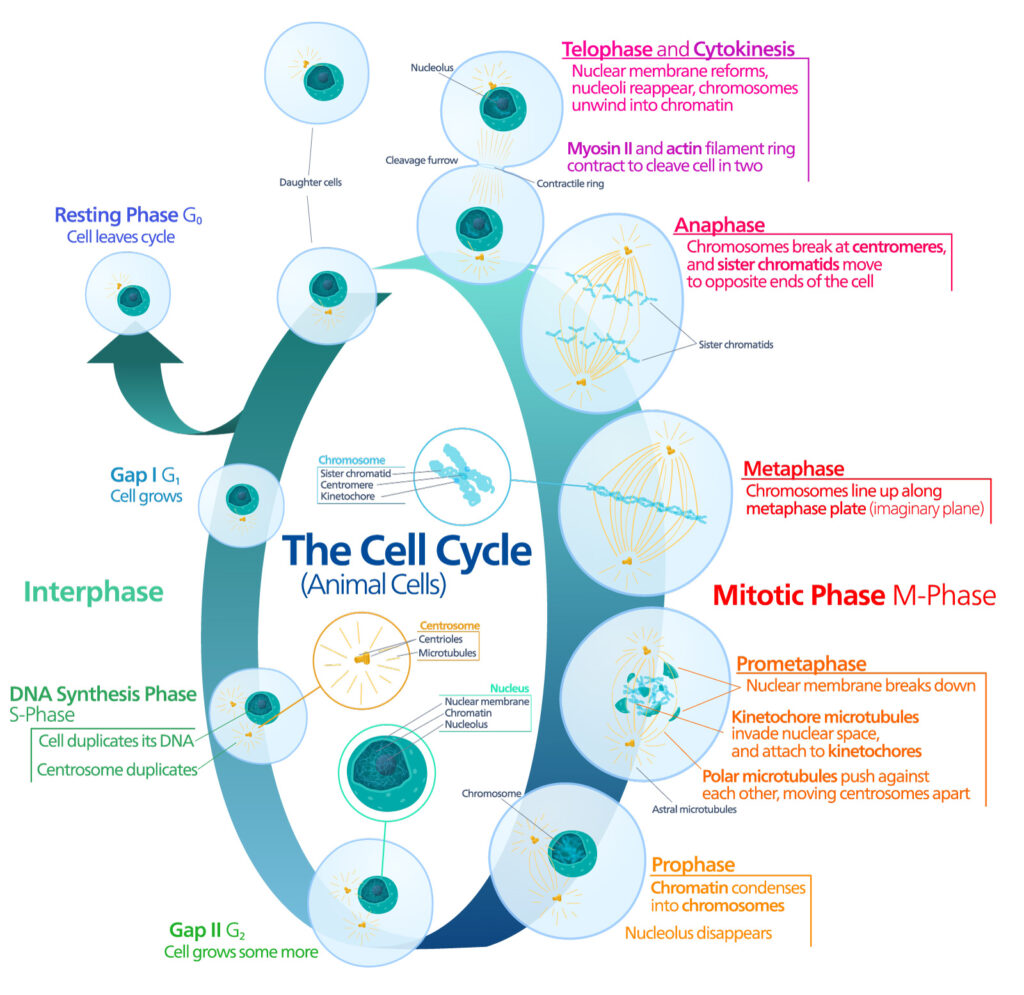
Pioneering work published in 1961 by Leonard Hayflick and Paul Moorhead challenged that theory (reviewed in 1). Their research showed that normal cells in culture have a finite capacity to replicate. After they reach a certain number of replicative cycles, cells stop dividing. Hayflick and Moorhead made the important distinction between normal human cells and cultured cancer cells, which are truly immortal. In later years, the limit to the number of replicative cycles normal human cells can undergo became known as the Hayflick limit. Although some scientists still express skepticism about these findings, the Hayflick limit is widely recognized as a fundamental principle of cell biology.
Glucose is an energy metabolite necessary for cellular survival and growth whether or not the cell is part of a tumor. Not only do cancer cells switch from oxidative phosphorylation to aerobic glycolysis (the Warburg effect) to gain more glucose, a hallmark of cancer, but they also increase the amount of glucose taken up from the surrounding extracellular space. However, the lack of glucose can have a negative effect on cells, causing them to become apoptotic in the absence of this metabolite. Cancer cells have methods to get around the requirement for glucose, including upregulating glucose transporters to improve access to the energy metabolite. In this Redox Biology article, researchers describe how activating androgen receptor in response to a lack of glucose affects the amount of GLUT1 expressed on prostate cancer cells, making the cells resistant to glucose deprivation.
To set the stage, two prostate cancer cell lines, LNCaP, an androgen-sensitive cell line, and LNCaP-R, an androgen-insensitive cell line, were deprived of glucose. Both cell lines showed signs of cell death, but LNCaP-R cells died in greater numbers. To probe how LNCaP cells died, several inhibitors (a pan-caspase inhibitor, two necroptosis inhibitors and a ferroptosis inhibitor) were added but did not change the way the cells died. However, an autophagy inhibitor enhanced cell death, suggesting the cells were necrotic not apoptotic. Teasing apart if the necrosis of LNCaP cells was due to glucose availability or merely disrupted glycolysis, the glucose analog 2DG was added to the medium with glucose. The cells survived when treated with 2DG, suggesting it was the absence of glucose that induced necrosis. When LNCaP cells were cultivated in medium that replaced glucose with mannose or fructose, the cells survived, another point in favor of sugar depletion causing cell death.
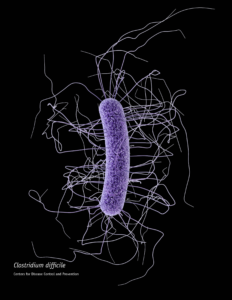
When someone is admitted to a hospital for an illness, the hope is that medical care and treatment will help them them feel better. However, nosocomial infections—infections acquired in a health-care setting—are becoming more prevalent and are associated with an increased mortality rate worldwide. This is largely due to the misuse of antibiotics, allowing some bacteria to become resistant. Furthermore, when an antibiotic wipes out the “good” bacteria that comprise the human microbiome, it leaves a patient vulnerable to opportunistic infections that take advantage of disruptions to the gut microbiota.
One such bacteria, Clostridium difficile, is of growing concern world-wide since it is resistant to many different antibiotics. When a patient is treated with an antibiotic, C. difficile can thrive in the intestinal tract without other bacteria populating the gut. C. difficile infection is the leading cause of antibiotic-associated diarrhea. While symptoms can be mild, aggressive infection can lead to pseudomembranous colitis—a severe inflammation of the colon which can be life-threatening.
C. difficile causes disease by releasing two large toxins, TcdA and TcdB. Understanding the role these toxins play in colonic disease is important for treatment strategies. However, most published research data only report the effects of the toxins independently. A 2016 study demonstrated a method of comparing the toxins side-by-side using the same time points and cell assays to investigate the role each toxin plays in the cell death that leads to disease of the colon. Continue reading “A Tale of Two Toxins: the mechanisms of cell death in Clostridium difficile infections”
In today’s post, guest blogger, Martha O’Brien, PhD, provides a preview of her upcoming AAI poster and block symposium talk on the inflammasome, caspase-1 activity and pyroptosis.
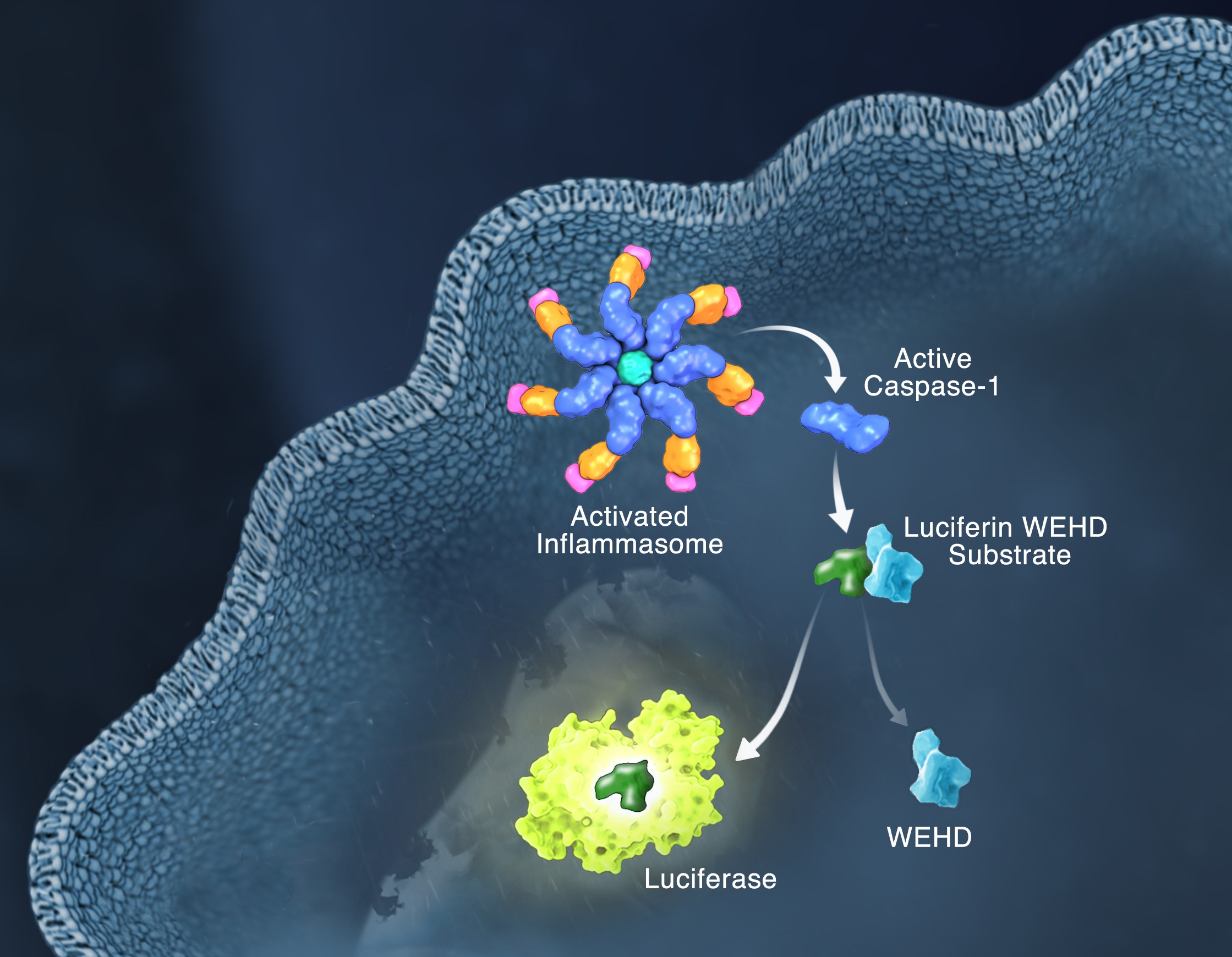
Schematic of the Caspase-Glo 1 Inflammasome Assay.
Responding rapidly to microbial pathogens and damage-associated molecular markers is critical to our innate immune system. Caspase-1 is pivotal in this process leading to processing and release of essential cytokines and an immunogenic form of cell death, termed pyroptosis. Upon sensing pathogen-associated and damage-associated molecular patterns (PAMPs and DAMPs), innate immune cells form inflammasome protein complexes that recruit and activate caspase-1 (canonical inflammasomes). In addition, other inflammatory caspases, 4 and 5 in humans and 11 in mice, directly bind bacterial lipopolysaccharides (LPS), triggering pyroptosis (non-canonical inflammasome). LPS-triggered non-canonical inflammasomes in mice and humans ultimately lead to canonical inflammasome engagement and caspase-1 activation (1–3). Caspase-1 was originally termed interleukin converting enzyme (ICE) for its well-established role in processing IL-1ß and IL-18, two important inflammation cytokines. How caspase-1 mediates pyroptosis is less well understood, but is beginning to be delineated. Recently, a substrate of the inflammatory caspases, gasdermin D, was identified and its processed fragment, gasdermin-N domain, was shown to be required for pyroptosis in non-canonical inflammasome circumstances (4, 5). The precise role of gasdermin D in canonical inflammasome-triggered pyroptosis is still under investigation. Linking inflammatory caspases directly to pyroptosis is a notable step in understanding the mechanism of this important form of cell death.
Pyroptosis is clearly one means of releasing processed IL-1ß and IL-18 from the cell. However depending on the cell type and stimulus, there is evidence for inflammasome engagement, caspase-1 activation, and release of IL-1ß in the absence of cell death (6, 7). On the flip-side there is also evidence for caspase-1 mediated pyroptosis that helps clear bacteria, independent of IL-1ß and IL-18 involvement (8). To enable further studies on the inflammasome and in particular, assessing the connections between caspase-1 activation, pyroptosis, and cytokine release, Promega developed a new tool to conveniently monitor caspase-1 activation, the Caspase-Glo® 1 Inflammasome Assay. This bioluminescent, plate-based assay is used to measure caspase-1 activity directly in cell cultures or to monitor released caspase-1 activity in culture medium from treated cells. This flexibility allows easy multiplexing to monitor all three outcomes of inflammasome stimulation; caspase-1 activity, pyroptosis, and release of IL-1ß and IL-18. Caspase-1 activation typically is monitored indirectly with western blots of processed caspase-1. Now the activity of the enzyme can be monitored directly, providing accurate information on temporal aspects of the inflammasome. The assay can be readily combined with real-time measures of cell death (e.g., CellTox™ Green Cytotoxicity Assay) and some of the culture medium can be removed for IL-1ß/IL-18 assessment, leaving the cells and remaining culture medium for caspase-1 activity measurements.
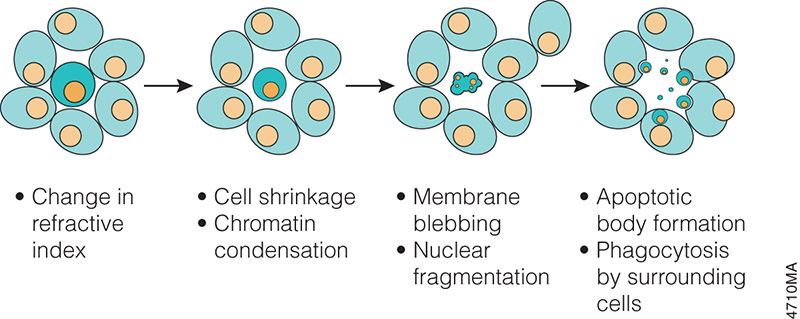
The concept of cell death as a normal cell fate was articulated only three years after Schleiden and Schwann introduced the Cell Theory when, in 1874, Vogt described natural cell death as an integral part of toad development (as cited in Cotter and Curtin, 2003). Since these early observations, natural cell death has been described “anew” several times. In 1885 Flemming provided the first morphological description of a natural cell death process, which we now label “apoptosis”, a term coined by Kerr and colleagues to describe the unique morphology associated with a cell death that differs from necrosis (as cited in Kerr et al. 1972).
Progression of morphology changes during apoptosis.
In the 1970s and 1980s, studies revealed that apoptosis not only had specific morphological characteristics but that it was also a tightly regulated process with specific biochemical characteristics. Studies of cell lineage in the nematode, Caenorhabditis elegans, showed that apoptosis was a normal feature of the nematode’s invariant developmental program (Hengartner, 1997). At the biochemical level, Wyllie showed that DNA degradation by a specific endonuclease during apoptosis resulted in a DNA ladder composed of mono- and oligonucleosomal-sized fragments (Wyllie, 1980).
These and many other studies have proven that apoptosis is a critical component of development, and when it doesn’t happen appropriately, it can be pathological, leading to cancers or other diseases. Therefore, understanding how and when apoptosis occurs and the many signals that can trigger this process is a focus of many laboratory experiments.
There are many ways to detect apoptosis in cells or tissues. This blog describes some of the most common ones.
XWe use cookies and similar technologies to make our website work, run analytics, improve our website, and show you personalized content and advertising. Some of these cookies are essential for our website to work. For others, we won’t set them unless you accept them. To learn more about our approach to Privacy we invite you to Read More
By clicking “Accept All”, you consent to the use of ALL the cookies. However you may visit Cookie Settings to provide a controlled consent.
We use cookies and similar technologies to make our website work, run analytics, improve our website, and show you personalized content and advertising. Some of these cookies are essential for our website to work. For others, we won’t set them unless you accept them. To find out more about cookies and how to manage cookies, read our Cookie Policy.
If you are located in the EEA, the United Kingdom, or Switzerland, you can change your settings at any time by clicking Manage Cookie Consent in the footer of our website.
Necessary cookies are absolutely essential for the website to function properly. These cookies ensure basic functionalities and security features of the website, anonymously.
Cookie
Duration
Description
cookielawinfo-checbox-analytics
11 months
This cookie is set by GDPR Cookie Consent plugin. The cookie is used to store the user consent for the cookies in the category "Analytics".
cookielawinfo-checbox-functional
11 months
The cookie is set by GDPR cookie consent to record the user consent for the cookies in the category "Functional".
cookielawinfo-checbox-others
11 months
This cookie is set by GDPR Cookie Consent plugin. The cookie is used to store the user consent for the cookies in the category "Other.
cookielawinfo-checkbox-advertisement
1 year
The cookie is set by GDPR cookie consent to record the user consent for the cookies in the category "Advertisement".
cookielawinfo-checkbox-necessary
11 months
This cookie is set by GDPR Cookie Consent plugin. The cookies is used to store the user consent for the cookies in the category "Necessary".
cookielawinfo-checkbox-performance
11 months
This cookie is set by GDPR Cookie Consent plugin. The cookie is used to store the user consent for the cookies in the category "Performance".
gdpr_status
6 months 2 days
This cookie is set by the provider Media.net. This cookie is used to check the status whether the user has accepted the cookie consent box. It also helps in not showing the cookie consent box upon re-entry to the website.
lang
This cookie is used to store the language preferences of a user to serve up content in that stored language the next time user visit the website.
viewed_cookie_policy
11 months
The cookie is set by the GDPR Cookie Consent plugin and is used to store whether or not user has consented to the use of cookies. It does not store any personal data.
Analytical cookies are used to understand how visitors interact with the website. These cookies help provide information on metrics the number of visitors, bounce rate, traffic source, etc.
Cookie
Duration
Description
SC_ANALYTICS_GLOBAL_COOKIE
10 years
This cookie is associated with Sitecore content and personalization. This cookie is used to identify the repeat visit from a single user. Sitecore will send a persistent session cookie to the web client.
vuid
2 years
This domain of this cookie is owned by Vimeo. This cookie is used by vimeo to collect tracking information. It sets a unique ID to embed videos to the website.
WMF-Last-Access
1 month 18 hours 24 minutes
This cookie is used to calculate unique devices accessing the website.
_ga
2 years
This cookie is installed by Google Analytics. The cookie is used to calculate visitor, session, campaign data and keep track of site usage for the site's analytics report. The cookies store information anonymously and assign a randomly generated number to identify unique visitors.
_gid
1 day
This cookie is installed by Google Analytics. The cookie is used to store information of how visitors use a website and helps in creating an analytics report of how the website is doing. The data collected including the number visitors, the source where they have come from, and the pages visted in an anonymous form.
Advertisement cookies are used to provide visitors with relevant ads and marketing campaigns. These cookies track visitors across websites and collect information to provide customized ads.
Cookie
Duration
Description
IDE
1 year 24 days
Used by Google DoubleClick and stores information about how the user uses the website and any other advertisement before visiting the website. This is used to present users with ads that are relevant to them according to the user profile.
test_cookie
15 minutes
This cookie is set by doubleclick.net. The purpose of the cookie is to determine if the user's browser supports cookies.
VISITOR_INFO1_LIVE
5 months 27 days
This cookie is set by Youtube. Used to track the information of the embedded YouTube videos on a website.
Performance cookies are used to understand and analyze the key performance indexes of the website which helps in delivering a better user experience for the visitors.
Cookie
Duration
Description
YSC
session
This cookies is set by Youtube and is used to track the views of embedded videos.
_gat_UA-62336821-1
1 minute
This is a pattern type cookie set by Google Analytics, where the pattern element on the name contains the unique identity number of the account or website it relates to. It appears to be a variation of the _gat cookie which is used to limit the amount of data recorded by Google on high traffic volume websites.
 When someone is admitted to a hospital for an illness, the hope is that medical care and treatment will help them them feel better. However, nosocomial infections—infections acquired in a health-care setting—are becoming more prevalent and are associated with an increased mortality rate worldwide. This is largely due to the misuse of antibiotics, allowing some bacteria to become resistant. Furthermore, when an antibiotic wipes out the “good” bacteria that comprise the human microbiome, it leaves a patient vulnerable to opportunistic infections that take advantage of disruptions to the gut microbiota.
When someone is admitted to a hospital for an illness, the hope is that medical care and treatment will help them them feel better. However, nosocomial infections—infections acquired in a health-care setting—are becoming more prevalent and are associated with an increased mortality rate worldwide. This is largely due to the misuse of antibiotics, allowing some bacteria to become resistant. Furthermore, when an antibiotic wipes out the “good” bacteria that comprise the human microbiome, it leaves a patient vulnerable to opportunistic infections that take advantage of disruptions to the gut microbiota.