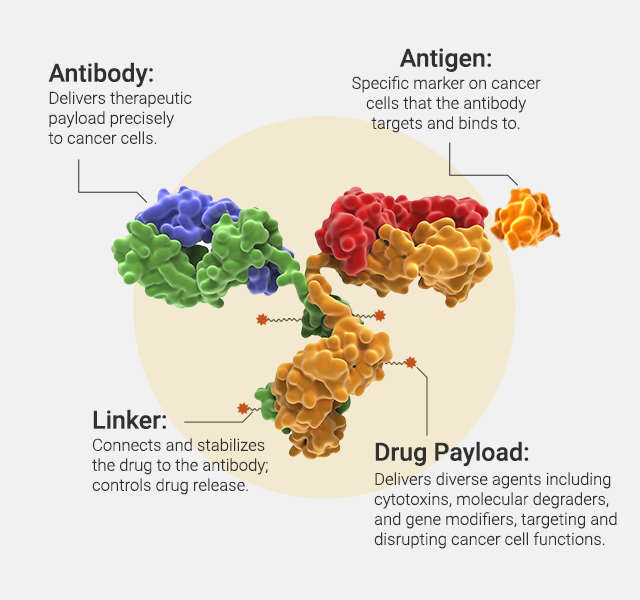
Antibody-drug conjugates (ADCs) are an increasingly powerful class of cancer therapeutics that combine the targeted precision of monoclonal antibodies with the cytotoxic potency of small-molecule drugs. By directing chemotherapy agents specifically to tumor cells, ADCs aim to maximize antitumor activity while minimizing damage to healthy tissues. One key challenge in ADC design is selecting the right target and payload—features that define efficacy, safety and resistance.
You have identified and cloned your protein of interest, but you want to explore its function. A protein fusion tag might help with your investigation. However, choosing a tag for your protein depends on what experiments you are planning. Do you want to purify the protein? Would you like to identify interacting proteins by performing pull-down assays? Are you interested in examining the endogenous biology of the protein? Here we cover the advantages and disadvantages of some protein tags to help you select the one that best suits your needs.
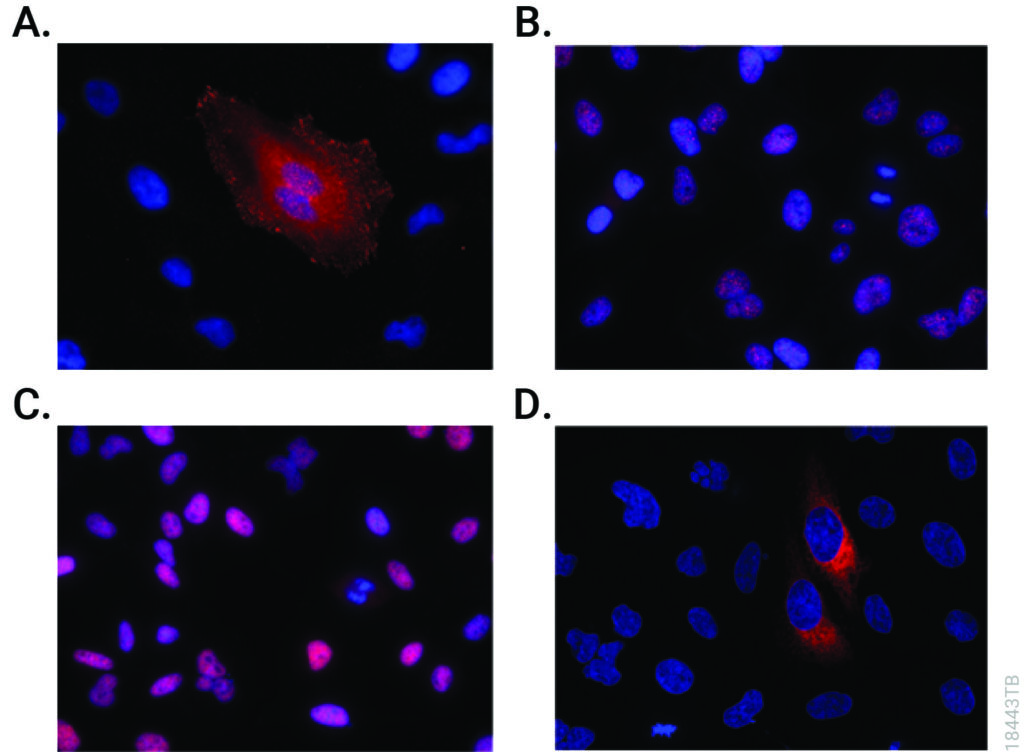
CRISPR-Cas9 editing knocked-in HiBiT at the endogenous locus of proteins with varying subcellular localization. Fixed CRISPR-modified clones or pools of cells were imaged by immunofluorescent staining using the Anti-HiBiT Monoclonal Antibody (red) and Hoechst dye (blue). Panel A. VCL-HiBiT pool. Panel B. SMARCA4-HiBiT clone. Panel C. HDAC2-HiBiT clone. Panel D. HSP90B1-HiBiT pool.
Affinity Tags
The most commonly used protein tags fall under the category of affinity tags. This means that the tag binds to another molecule or metal ion, making it easy to purify or pull down your protein of interest. In all cases, the tag will be fused to your protein of interest at either the amino (N) or carboxy (C) terminus by cloning into an expression vector. This protein fusion can then be expressed in cells or cell-free systems, depending on the promoter the vector contains.
Cell-free protein expression is a simplified and accelerated avenue for the transcription and/or translation of a specific protein in a quasi cell environment. An alternative to slower, more cumbersome cell-based methods, cell-free protein expression methods are simple and fast and can overcome toxicity and solubility issues sometimes experienced in the traditional E. coli expression systems.
Cell-free protein expression offers significant time savings over cell-based expression methods.
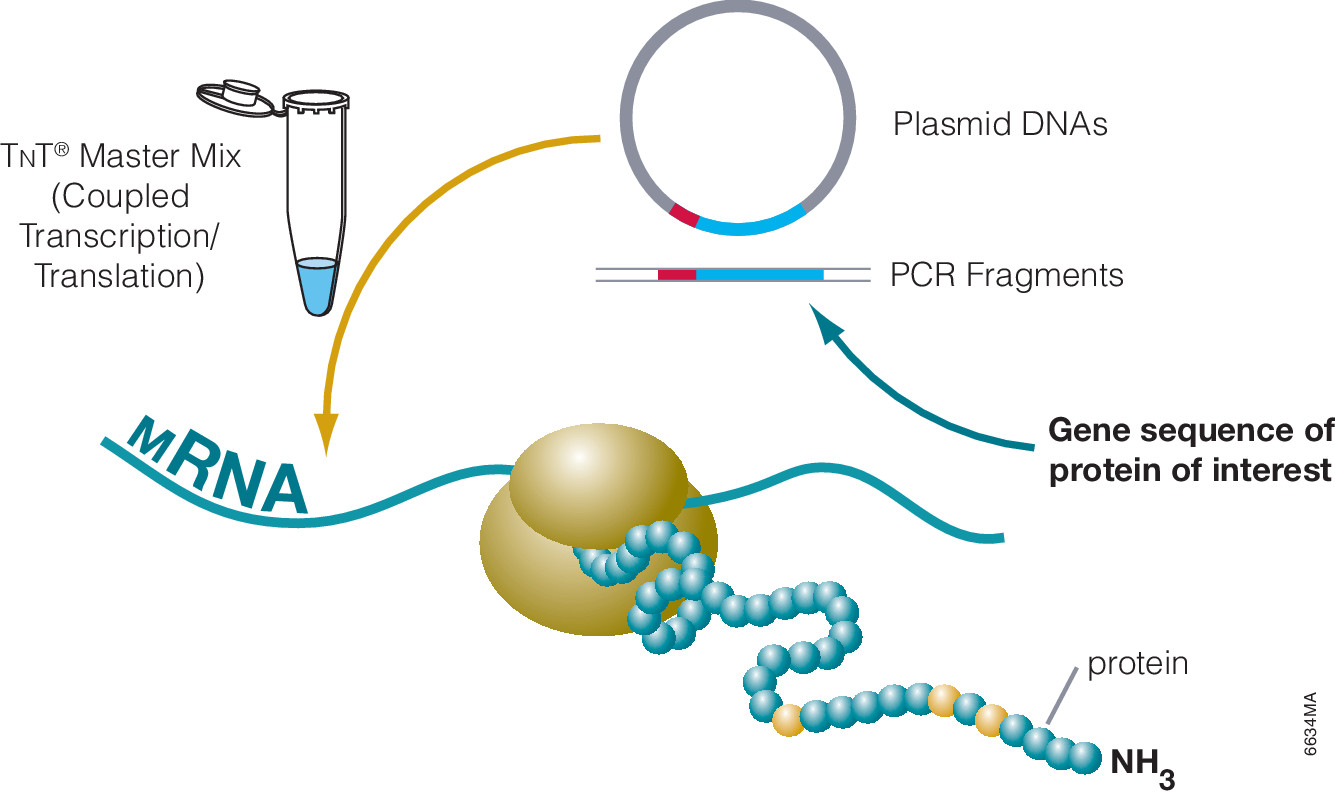
Cell-free protein expression offers a convenient method for generating small amounts of protein for a variety of applications (e.g., protein:protein interactions, protein: nucleic acid interactions, structural analysis, functional assays and toxicity screening). This approach lends itself to specific protein labeling with fluorescence, biotin, radioactivity or heavy atoms, via modified charged tRNA’s or amino acids. Cell-free protein expression systems provide quick access to proteins of interest and remain a staple in the collection of tools available for the elucidation of protein structure and function, understanding cellular pathways and mechanisms and high-throughput screening of compounds for drug discovery. There are a number of different cell-free expressions systems, each with different strengths. Deciding which one is right for you depends upon your research needs and goals.
Cell-free protein expression is a simplified and accelerated avenue for the transcription and/or translation of a specific protein in a quasi cell environment. An alternative to slower, more cumbersome cell-based methods, cell-free protein expression methods are simple and fast and can overcome toxicity and solubility issues sometimes experienced in traditional E. coli expression systems. Continue reading “Cell-free Expression: A System for Every Need”
Many applications require amounts of protein that cannot be obtained using a eukaryotic cell-free expression system. As an alternative, a prokaryotic system can be used when this need arises. The E. coli S30 T7 High-Yield Protein Expression System is designed to express up to 500μg/ml of protein in 1 hour from plasmid vectors containing a T7 promoter and a ribosome binding site. The protein expression system provides an extract that contains T7 RNA polymerase for transcription and is deficient in OmpT endoproteinase and lon protease activity. All other necessary components in the system are optimized for protein expression. This results in greater stability and enhanced expression of target proteins.The following references highlight the use of this system for a variety of unique applications:
mRNAs commonly exhibit differing salt requirements for optimal translation. Small variations in salt concentration can lead to dramatic differences in translation efficiency. The Flexi® Rabbit Reticulocyte Lysate System allows translation reactions to be optimized for a wide range of parameters, including Mg2+ and K+ concentrations and the choice of adding DTT. To help optimize Mg2+ for a specific message, the endogenous Mg2+ concentration of each lysate batch is stated in the product information included with this product.
The following references utilize the features of Flexi Rabbit Reticulocyte Lysate System to investigate certain parameters of translation:
Sato, V. et al. (2007) Measles virus N protein inhibits host translation by binding to eIF3-p40. J. Vir.81, 11569–76. Charaterized the effect of the measles virus N protein binding to the translation initiation factor eIF3-p40 on the expression of various proteins in rabbit reticulocyte lysate.
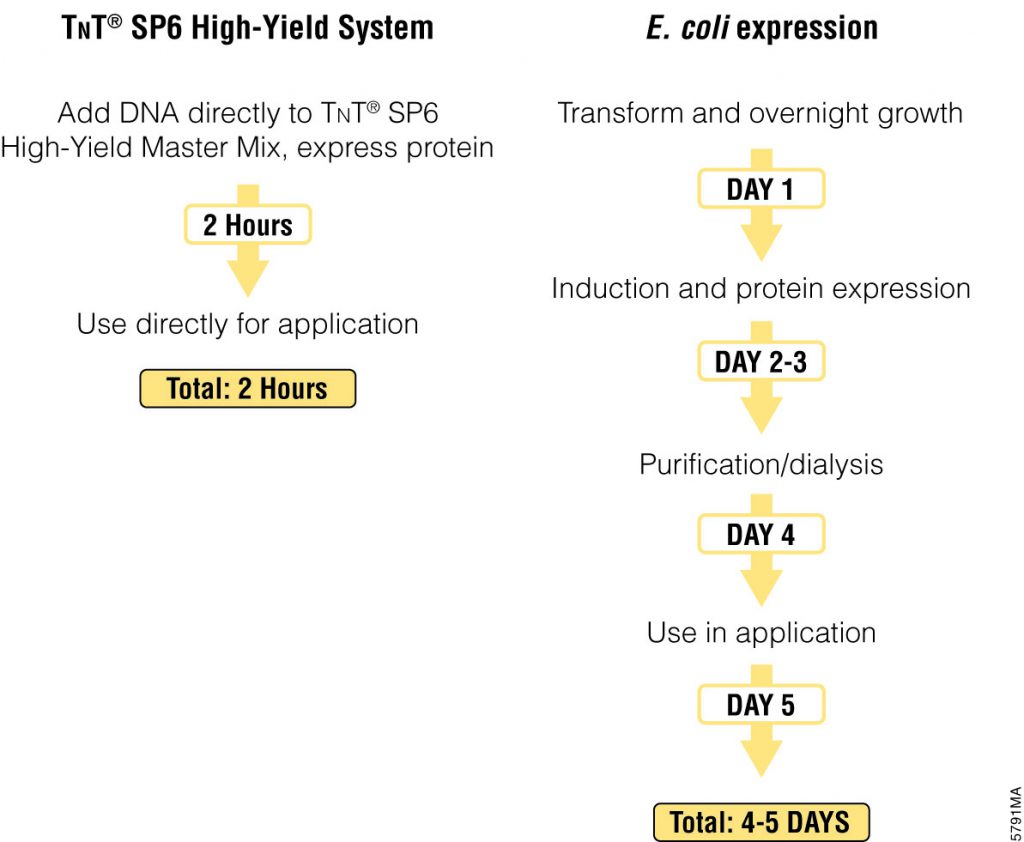
The TNT® SP6 High-Yield Protein Expression System uses a high-yield wheat germ extract supplemented with SP6 RNA polymerase and other components. Coupling transcriptional and translational activities eliminates the inconvenience of separate in vitro transcription and purification steps for the mRNA, while maintaining the high levels of protein expression. All that is required is the addition of DNA templates containing the SP6 promoter and the protein coding region for the protein of interest. Furthermore no specialized equipment is required for protein screening and production. The system enables the expression of approximately 100µg/ml of protein in batch reaction and 200–440µg/ml in dialysis reaction in 10–20 hours .
Zhao, L. et.al. (2010) J. Struct. Genomics11, 201–9, the Northeast Structural Genomics Consortium (www.nesg.org) in their quest to express 5,000 eukaryotic proteins, report that even with different cloning strategies they could only produce 26% of the proteins in a soluble form. To improve the efficiency of expressing soluble protein, they investigated the use of wheat germ cell free system as a alternative to E.coli.
In this publication 59 human constructs were expressed in both E.coli and the wheat germ cell free system. Only 30% of human proteins could be produced in a soluble form using E.coli -based expression. Some 70% could be produced using the TNT® SP6 High Yield Wheat Germ system.
To further demonstrate the utility of expressing proteins that were suitable for structural studies from a wheat germ-based system, two of the proteins were isotope enriched and analyzed successfully by 2D NMR.
The analysis of functional protein typically requires lengthy laborious cell based protein expression that can be complicated by the lack of stability or solubility of the purified protein. Cell free protein expression eliminates the requirement for cell culture thus providing quick access to the protein of interest (1).
The HaloTag® Technology provides efficient, covalent and oriented protein immobilization of the fusion protein to solid surfaces (2).
Leippe et al. demonstrated the feasibility of using cell free expression and the HaloTag technology to express and capture a fusion protein for the rapid screening of protein kinase activity (3). The catalytic subunit of human cAMP dependent protein kinase was expressed in a variety of cell free expression formats as a HaloTag fusion protein. The immobilized cPKA fusion protein was assayed directly on magnetic beads in the active form and was shown to be inhibited by known PKA inhibitory compounds.
Therefore this unique combination of protein expression and capture technologies can greatly facilitate the process of activity screening and characterization of potential inhibitors
Pull-down assays probe interactions between a protein of interest that is expressed as fusion protein (e.g., (e.g., bait) and the potential interacting partners (prey).
In a pull-down assay one protein partner is expressed as a fusion protein (e.g., bait protein) in E. coli and then immobilized using an affinity ligand specific for the fusion tag. The immobilized
bait protein can then be incubated with the prey protein. The source of the prey protein depends on whether the experiment is designed to confirm an interaction or to identify new interactions. After a series of wash steps, the entire complex can be eluted from the affinity support using SDS-PAGE loading buffer or by competitive analyte elution, then evaluated by SDS-PAGE.
Successful interactions can be detected by Western blotting with specific antibodies to both the prey and bait proteins, or measurement of radioactivity from a [35S] prey protein. bait) and potential interacting partners (prey).
The most commonly used method to generate a bait protein is expression as a fusion protein contain a GST (glutathione-S transferase) tag in E. coli. This is followed by immobilization on particles that contain reduced glutathione, which binds to the GST tag of the fusion protein. The primary advantage of a GST tag is that it can increase the solubility of insoluble or semi-soluble proteins expressed in E. coli.
Among fusion tags, His-tag is the most widely used and has several advantages including: 1) It’s small in size, which renders it less immunogenically active, and often it does not need to be removed from the purified protein for downstream applications; 2) There are a large number of commercial vectors available for expressing His-tagged proteins; 3) The tag may be placed at either the N or C terminus; 4) The interaction of the His-tag does not depend on the tag structure, making it possible to purify otherwise insoluble proteins using denaturing conditions.
Cell-free protein synthesis has emerged as powerful alternative to cell based protein expression for functional and structural proteomics. The TNT® SP6 High-Yield Protein Expression System uses a high-yield wheat germ extract supplemented with SP6 RNA polymerase and other components. Coupling transcriptionaland translational activities eliminates the inconvenience of separate in vitro transcription and purification steps for the mRNA, while maintaining the high levels of protein expression (1).
XWe use cookies and similar technologies to make our website work, run analytics, improve our website, and show you personalized content and advertising. Some of these cookies are essential for our website to work. For others, we won’t set them unless you accept them. To learn more about our approach to Privacy we invite you to Read More
By clicking “Accept All”, you consent to the use of ALL the cookies. However you may visit Cookie Settings to provide a controlled consent.
We use cookies and similar technologies to make our website work, run analytics, improve our website, and show you personalized content and advertising. Some of these cookies are essential for our website to work. For others, we won’t set them unless you accept them. To find out more about cookies and how to manage cookies, read our Cookie Policy.
If you are located in the EEA, the United Kingdom, or Switzerland, you can change your settings at any time by clicking Manage Cookie Consent in the footer of our website.
Necessary cookies are absolutely essential for the website to function properly. These cookies ensure basic functionalities and security features of the website, anonymously.
Cookie
Duration
Description
cookielawinfo-checbox-analytics
11 months
This cookie is set by GDPR Cookie Consent plugin. The cookie is used to store the user consent for the cookies in the category "Analytics".
cookielawinfo-checbox-functional
11 months
The cookie is set by GDPR cookie consent to record the user consent for the cookies in the category "Functional".
cookielawinfo-checbox-others
11 months
This cookie is set by GDPR Cookie Consent plugin. The cookie is used to store the user consent for the cookies in the category "Other.
cookielawinfo-checkbox-advertisement
1 year
The cookie is set by GDPR cookie consent to record the user consent for the cookies in the category "Advertisement".
cookielawinfo-checkbox-necessary
11 months
This cookie is set by GDPR Cookie Consent plugin. The cookies is used to store the user consent for the cookies in the category "Necessary".
cookielawinfo-checkbox-performance
11 months
This cookie is set by GDPR Cookie Consent plugin. The cookie is used to store the user consent for the cookies in the category "Performance".
gdpr_status
6 months 2 days
This cookie is set by the provider Media.net. This cookie is used to check the status whether the user has accepted the cookie consent box. It also helps in not showing the cookie consent box upon re-entry to the website.
lang
This cookie is used to store the language preferences of a user to serve up content in that stored language the next time user visit the website.
viewed_cookie_policy
11 months
The cookie is set by the GDPR Cookie Consent plugin and is used to store whether or not user has consented to the use of cookies. It does not store any personal data.
Analytical cookies are used to understand how visitors interact with the website. These cookies help provide information on metrics the number of visitors, bounce rate, traffic source, etc.
Cookie
Duration
Description
SC_ANALYTICS_GLOBAL_COOKIE
10 years
This cookie is associated with Sitecore content and personalization. This cookie is used to identify the repeat visit from a single user. Sitecore will send a persistent session cookie to the web client.
vuid
2 years
This domain of this cookie is owned by Vimeo. This cookie is used by vimeo to collect tracking information. It sets a unique ID to embed videos to the website.
WMF-Last-Access
1 month 18 hours 24 minutes
This cookie is used to calculate unique devices accessing the website.
_ga
2 years
This cookie is installed by Google Analytics. The cookie is used to calculate visitor, session, campaign data and keep track of site usage for the site's analytics report. The cookies store information anonymously and assign a randomly generated number to identify unique visitors.
_gid
1 day
This cookie is installed by Google Analytics. The cookie is used to store information of how visitors use a website and helps in creating an analytics report of how the website is doing. The data collected including the number visitors, the source where they have come from, and the pages visted in an anonymous form.
Advertisement cookies are used to provide visitors with relevant ads and marketing campaigns. These cookies track visitors across websites and collect information to provide customized ads.
Cookie
Duration
Description
IDE
1 year 24 days
Used by Google DoubleClick and stores information about how the user uses the website and any other advertisement before visiting the website. This is used to present users with ads that are relevant to them according to the user profile.
test_cookie
15 minutes
This cookie is set by doubleclick.net. The purpose of the cookie is to determine if the user's browser supports cookies.
VISITOR_INFO1_LIVE
5 months 27 days
This cookie is set by Youtube. Used to track the information of the embedded YouTube videos on a website.
Performance cookies are used to understand and analyze the key performance indexes of the website which helps in delivering a better user experience for the visitors.
Cookie
Duration
Description
YSC
session
This cookies is set by Youtube and is used to track the views of embedded videos.
_gat_UA-62336821-1
1 minute
This is a pattern type cookie set by Google Analytics, where the pattern element on the name contains the unique identity number of the account or website it relates to. It appears to be a variation of the _gat cookie which is used to limit the amount of data recorded by Google on high traffic volume websites.