In recent years, scientists have been hot on the trail of transcription factor FOXO3, tracing its involvement in various tumor-centric activities comprising the many trademarks of cancer, from drug resistance to metastasis to tumor angiogenesis.
FOXO3 is a member of the O sub-class of the forkhead box family of transcription factors. The forkhead box (FOX) family is characterized by a fork head DNA-binding domain (DBD), comprised of around 100 amino acids. They have also proven themselves to be a family of many hats, functioning in diverse roles ranging from metabolism, immunology, cell-cycle control, development, as well as cancer (1). The forkhead box O (FOXO) sub-class alone has demonstrated involvement in a variety of cellular outcomes, from drug resistance and longevity to apoptosis induction.
Due to its pro-apoptotic and anti-proliferative proclivity, FOXO3 has been previously identified as a tumor suppressor gene. However, more and more studies have begun to flip the narrative on FOXO3, portraying it more as a devoted henchman, due to its roles in drug and radiotherapy resistance, cell-cycle arrest and long-term maintenance of leukemia-initiating stem cells in a variety of cancer types, including breast cancer, pancreatic cancer, glioblastoma, and both acute and chronic myeloid leukemia.
In today’s post, guest blogger, Martha O’Brien, PhD, provides a preview of her upcoming AAI poster and block symposium talk on the inflammasome, caspase-1 activity and pyroptosis.
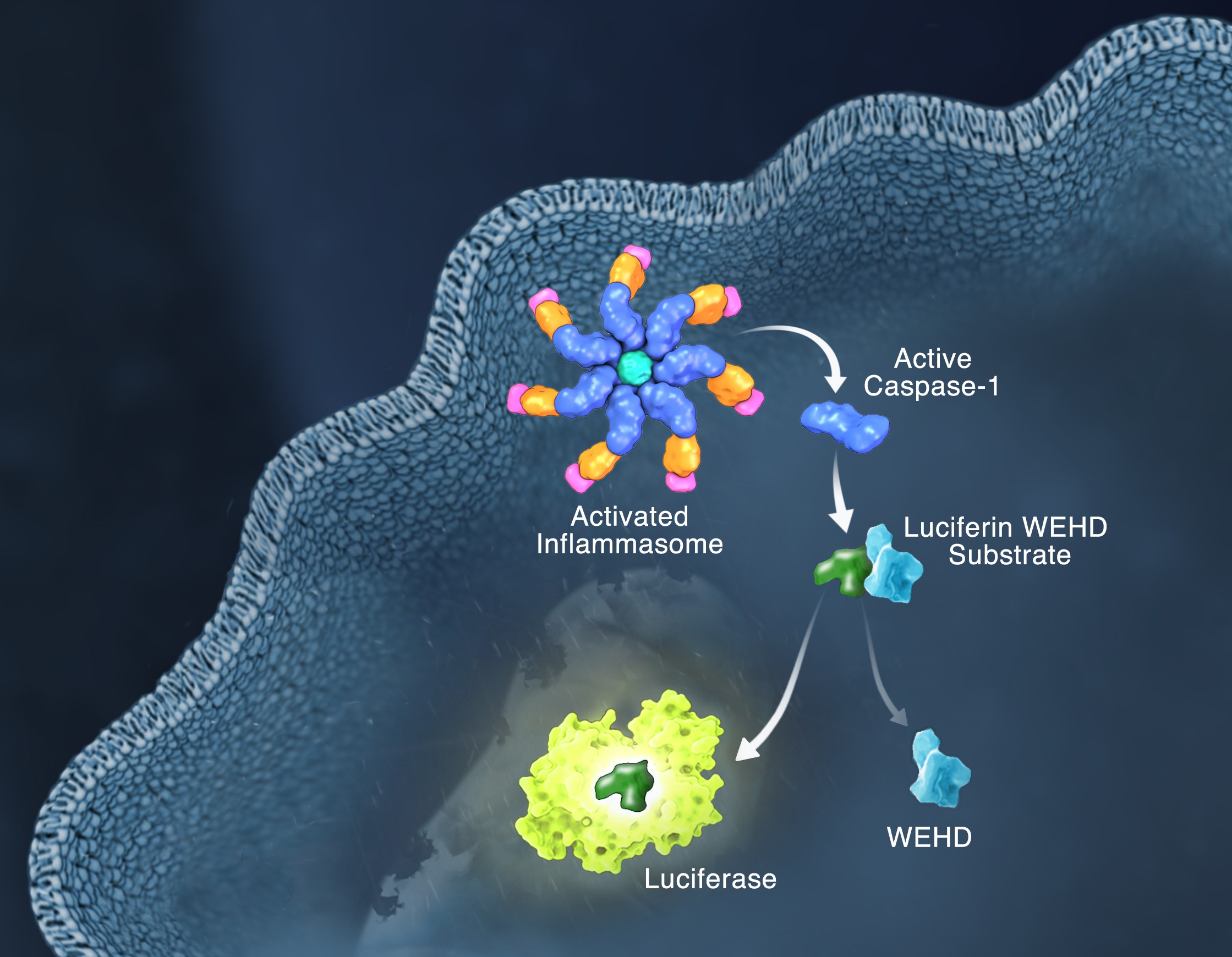
Schematic of the Caspase-Glo 1 Inflammasome Assay.
Responding rapidly to microbial pathogens and damage-associated molecular markers is critical to our innate immune system. Caspase-1 is pivotal in this process leading to processing and release of essential cytokines and an immunogenic form of cell death, termed pyroptosis. Upon sensing pathogen-associated and damage-associated molecular patterns (PAMPs and DAMPs), innate immune cells form inflammasome protein complexes that recruit and activate caspase-1 (canonical inflammasomes). In addition, other inflammatory caspases, 4 and 5 in humans and 11 in mice, directly bind bacterial lipopolysaccharides (LPS), triggering pyroptosis (non-canonical inflammasome). LPS-triggered non-canonical inflammasomes in mice and humans ultimately lead to canonical inflammasome engagement and caspase-1 activation (1–3). Caspase-1 was originally termed interleukin converting enzyme (ICE) for its well-established role in processing IL-1ß and IL-18, two important inflammation cytokines. How caspase-1 mediates pyroptosis is less well understood, but is beginning to be delineated. Recently, a substrate of the inflammatory caspases, gasdermin D, was identified and its processed fragment, gasdermin-N domain, was shown to be required for pyroptosis in non-canonical inflammasome circumstances (4, 5). The precise role of gasdermin D in canonical inflammasome-triggered pyroptosis is still under investigation. Linking inflammatory caspases directly to pyroptosis is a notable step in understanding the mechanism of this important form of cell death.
Pyroptosis is clearly one means of releasing processed IL-1ß and IL-18 from the cell. However depending on the cell type and stimulus, there is evidence for inflammasome engagement, caspase-1 activation, and release of IL-1ß in the absence of cell death (6, 7). On the flip-side there is also evidence for caspase-1 mediated pyroptosis that helps clear bacteria, independent of IL-1ß and IL-18 involvement (8). To enable further studies on the inflammasome and in particular, assessing the connections between caspase-1 activation, pyroptosis, and cytokine release, Promega developed a new tool to conveniently monitor caspase-1 activation, the Caspase-Glo® 1 Inflammasome Assay. This bioluminescent, plate-based assay is used to measure caspase-1 activity directly in cell cultures or to monitor released caspase-1 activity in culture medium from treated cells. This flexibility allows easy multiplexing to monitor all three outcomes of inflammasome stimulation; caspase-1 activity, pyroptosis, and release of IL-1ß and IL-18. Caspase-1 activation typically is monitored indirectly with western blots of processed caspase-1. Now the activity of the enzyme can be monitored directly, providing accurate information on temporal aspects of the inflammasome. The assay can be readily combined with real-time measures of cell death (e.g., CellTox™ Green Cytotoxicity Assay) and some of the culture medium can be removed for IL-1ß/IL-18 assessment, leaving the cells and remaining culture medium for caspase-1 activity measurements.
XWe use cookies and similar technologies to make our website work, run analytics, improve our website, and show you personalized content and advertising. Some of these cookies are essential for our website to work. For others, we won’t set them unless you accept them. To learn more about our approach to Privacy we invite you to Read More
By clicking “Accept All”, you consent to the use of ALL the cookies. However you may visit Cookie Settings to provide a controlled consent.
We use cookies and similar technologies to make our website work, run analytics, improve our website, and show you personalized content and advertising. Some of these cookies are essential for our website to work. For others, we won’t set them unless you accept them. To find out more about cookies and how to manage cookies, read our Cookie Policy.
If you are located in the EEA, the United Kingdom, or Switzerland, you can change your settings at any time by clicking Manage Cookie Consent in the footer of our website.
Necessary cookies are absolutely essential for the website to function properly. These cookies ensure basic functionalities and security features of the website, anonymously.
Cookie
Duration
Description
cookielawinfo-checbox-analytics
11 months
This cookie is set by GDPR Cookie Consent plugin. The cookie is used to store the user consent for the cookies in the category "Analytics".
cookielawinfo-checbox-functional
11 months
The cookie is set by GDPR cookie consent to record the user consent for the cookies in the category "Functional".
cookielawinfo-checbox-others
11 months
This cookie is set by GDPR Cookie Consent plugin. The cookie is used to store the user consent for the cookies in the category "Other.
cookielawinfo-checkbox-advertisement
1 year
The cookie is set by GDPR cookie consent to record the user consent for the cookies in the category "Advertisement".
cookielawinfo-checkbox-necessary
11 months
This cookie is set by GDPR Cookie Consent plugin. The cookies is used to store the user consent for the cookies in the category "Necessary".
cookielawinfo-checkbox-performance
11 months
This cookie is set by GDPR Cookie Consent plugin. The cookie is used to store the user consent for the cookies in the category "Performance".
gdpr_status
6 months 2 days
This cookie is set by the provider Media.net. This cookie is used to check the status whether the user has accepted the cookie consent box. It also helps in not showing the cookie consent box upon re-entry to the website.
lang
This cookie is used to store the language preferences of a user to serve up content in that stored language the next time user visit the website.
viewed_cookie_policy
11 months
The cookie is set by the GDPR Cookie Consent plugin and is used to store whether or not user has consented to the use of cookies. It does not store any personal data.
Analytical cookies are used to understand how visitors interact with the website. These cookies help provide information on metrics the number of visitors, bounce rate, traffic source, etc.
Cookie
Duration
Description
SC_ANALYTICS_GLOBAL_COOKIE
10 years
This cookie is associated with Sitecore content and personalization. This cookie is used to identify the repeat visit from a single user. Sitecore will send a persistent session cookie to the web client.
vuid
2 years
This domain of this cookie is owned by Vimeo. This cookie is used by vimeo to collect tracking information. It sets a unique ID to embed videos to the website.
WMF-Last-Access
1 month 18 hours 24 minutes
This cookie is used to calculate unique devices accessing the website.
_ga
2 years
This cookie is installed by Google Analytics. The cookie is used to calculate visitor, session, campaign data and keep track of site usage for the site's analytics report. The cookies store information anonymously and assign a randomly generated number to identify unique visitors.
_gid
1 day
This cookie is installed by Google Analytics. The cookie is used to store information of how visitors use a website and helps in creating an analytics report of how the website is doing. The data collected including the number visitors, the source where they have come from, and the pages visted in an anonymous form.
Advertisement cookies are used to provide visitors with relevant ads and marketing campaigns. These cookies track visitors across websites and collect information to provide customized ads.
Cookie
Duration
Description
IDE
1 year 24 days
Used by Google DoubleClick and stores information about how the user uses the website and any other advertisement before visiting the website. This is used to present users with ads that are relevant to them according to the user profile.
test_cookie
15 minutes
This cookie is set by doubleclick.net. The purpose of the cookie is to determine if the user's browser supports cookies.
VISITOR_INFO1_LIVE
5 months 27 days
This cookie is set by Youtube. Used to track the information of the embedded YouTube videos on a website.
Performance cookies are used to understand and analyze the key performance indexes of the website which helps in delivering a better user experience for the visitors.
Cookie
Duration
Description
YSC
session
This cookies is set by Youtube and is used to track the views of embedded videos.
_gat_UA-62336821-1
1 minute
This is a pattern type cookie set by Google Analytics, where the pattern element on the name contains the unique identity number of the account or website it relates to. It appears to be a variation of the _gat cookie which is used to limit the amount of data recorded by Google on high traffic volume websites.